| 1. | ||
| 2. | ||
| 3. | ||
| 4. | ||
| 5. | ||
Landmark-Based Geometric Mophometric Analysis on Nepenthes saranganiensis Kurata in Relationship to the Plant’s Trapping Strategies
John Vincent P. Anino II, Dave P. Buenavista*
Department of Biology, Central Mindanao University, University Town, Musuan, Bukidnon, Philippines
Email address
 (J. V. P. Anino II)
(J. V. P. Anino II)  (D. P. Buenavista)
(D. P. Buenavista) Citation
John Vincent P. Anino II, Dave P. Buenavista. Landmark-Based Geometric Mophometric Analysis on Nepenthes saranganiensis Kurata in Relationship to the Plant’s Trapping Strategies. International Journal of Bioinformatics and Computational Biology. Vol. 1, No. 1, 2016, pp. 1-8.
Abstract
This study was conducted to determine the possible trapping strategies of N. saranganiensis Kurata through landmark-based geometric morphometric analysis. Specifically, it aimed to: (a) determine the quantitative geometric shapes in the upper pitcher peristome and body of N. saranganiensis and (b) determine the ecological traits of the selected wild populations of Nepenthes. High quality photographs of the trap geometry of the plant were taken i.e., the peristome and body shape. The peristome and body shape were studied using a landmark-based methodology that eliminates the effect of variation in the location, orientation, and scale of the specimens. Geographical coordinates, elevation, habitat, air temperature, relative humidity, and animal associates were considered to assess the plant’s ecological traits. Based from the Relative Warps Analysis on the shapes of the peristome and pitcher body of N. saranganiensis, results revealed that the most variable part of the peristome were the lateral and basal regions including the entire structure of the pitcher from the slippery zone, transition zone and digestive zone. Histogram distributions have shown that most of the specimens tend to have wider peristomes and bodies when compared to the shape consensus. Putative functional morphology based from the analysis showed that morphological features of N. saranganiensis exhibits a prominent and developed slippery zone and narrow peristome in relation to its body size as an adaptation for "Dry Syndrome" trapping strategy as this species mainly specialized in prey retention carnivory.
Keywords
Functional Morphology, Trapping Strategy, Nepenthes, Geometric Morphometrics, Dry Syndrome
1. Introduction
Nepenthes L. is a genus of carnivorous plants belonging to the order Caryophyllales [1]. Approximately 140 species are known globally, 38 species of which are found in the Philippines particularly in the island of Mindanao [2]. Carnivorous plants are excellent specimens on studying the evolution of functional morphology in plants in which they use specialized leaves to trap and provide essential nutrients which enable these plants to inhabit nutrient-poor habitats [3-5].
There have been growing number of evidences supporting that Nepenthes pitcher plants have evolved several alternative trapping strategies in which this specializations may have been the main driving force towards the speciation and rapid diversification of the genus [7-15].
However, the methods of Bauer et al. [3], Benz et al. [22] and Moran et al. [8] only included the measurement of peristome width, downward angle of the peristome and the length of the pitcher body and not the shape of the entire pitcher and some of the data gathered were only based on plants grown in different herbaria. Morphological variations in pitcher plants were previously reported to be influenced primarily by geographical locations, climatic conditions and environmental conditions [37]. In addition to that, plant or pitcher size and color variations are considered as minor and /or unstable morphological characteristics and thus considered insignificant [23]. In addition, N. saranganiensis Kurata is considered to be one of the most poorly understood Philippine endemic Nepenthes.
Geometric morphometrics has proven to be a powerful tool in resolving taxonomic problems as well as understanding the evolutionary biology of certain species of plants and animals at the species level but also can be utilized to describe the differences of the shape within a single species such as describing the mandible shapes of Diacamma (Formicidae) [24], sea snail shell shapes of Terebralia [28], head shapes of Lepidopteran Ostrinia larvae [29] and describing and comparing plant shapes between species such as Dactylorhiza [17], Quercus [31], Ophioglossum [25] and few other groups in which had also provided significant quantitative results in resolving some taxonomic problems.
Due to the lack of information and data pertaining to the functional morphology of Nepenthes pitchers, this study was carried out with the assumption that geometric morphometric analysis can be utilized on investigating the trapping strategies of certain less understood species of Nepenthes in conjunction to the habitat description where these plants originally are found.
2. Methodology
The study was conducted in Mt. Kalatungan Range, it is located in the north central portion of Pangantucan, Bukidon, Philippines (Fig. 1).
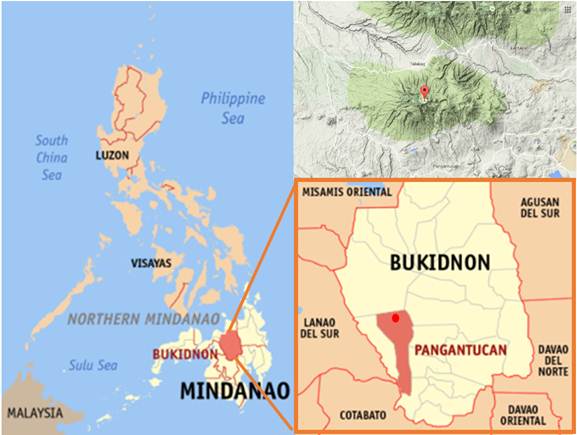
Fig. 1. Map showing Mount Kalatungan Range, Bukidnon, Philippines.
Since the upper pitcher presents significant taxonomic value, at least three representatives of the upper pitchers were selected from an observed clumping populations. Elevation and geographical coordinates were measured using GPS and recorded in each of the sampled population. High quality photographs of the trap geometry of the plant were taken i.e., the peristome and body shape. The pitcher peristome and body were photographed 90° from the ground. Peristome and body shape was studied using a landmark-based methodology that eliminates the effect of variation in the location, orientation, and scale of the specimens. A total of 48 anatomical and mathematical landmarks were assigned along the upper pitcher peristome (Fig. 2) and 69 anatomical and mathematical landmarks were assigned along the outline of pitcher body (Fig. 3) using an image analysis and processing software TpsDig freeware 2.12 [32]. The two-dimensional coordinates of these landmarks were determined for each Nepenthes specimen. Then the generalized orthogonal least squares Procrustes average configuration of landmarks was computed using generalized Procustes Analysis (GPA) superimposition method. GPA was performed using software tpsRelw, ver. 1.46 [33]. Histogram and box plots were generated using PAST software from the relative warps of the shell shapes. Histograms are a powerful display for comparing distributions. They provide a compact view of where the data are centered and how they are distributed over the range of the variable.

Fig. 2. Image showing 48 anatomical and mathematical landmarks assigned along the upper pitcher peristome.

Fig. 3. Image showing the 69 anatomical and mathematical landmarks assigned along the outline of the pitcher body.
Geographical coordinates were recorded using a GPS device. Spatial and temporal data such as air temperature and relative humidity were recorded with a thermometer for the former and a sling psychrometer, respectively. Habitat description, type locality and animal associates were determined and recorded on a field notebook. Results from the Landmark-based Geometric Morphometric analysis were used to determine and investigate the putative functional morphology of the N. saranganiensis.
3. Results and Observations
Relative warp analysis describes the variations observed in N. saranganiensis. This is presented in Table 1 and 2 and graphically illustrated in Fig. 6 and 7.

Fig. 4. Habitat of N. saranganiensis in the canopy mossy forest in Mt. Kalatungan, Pangantucan, Bukidnon Philippines.

Fig. 5. Arboreal Philautus sp. living inside the trap of N. saranganiensis.
N. saranganiensis was found in a mossy forest dominated by gymnosperms along with some tree ferns in Mt. Kalatungan, Pangantucan, Bukidnon, Philippines (fig. 4) with an elevation of 2020 masl and a corresponding geographical coordinates of 07°56.369’ N and 124°51.676’ E. The epiphytic plant was attached to the upper canopy and mid canopy of Phyllocladus sp. The recorded air temperature in the area was 17°C with a relative humidity of 90.5% which means that the area has a high saturation of moisture. It has also been confirmed that bryophytes contribute to the total humidity of a mossy environment [34]. The slippery zone N. saranganiensis is very distinguishable with its dull and matte white color that is visible in the inner surface of the pitcher (Fig. 2). The pitcher fluid was already highly diluted with rainwater and its viscosity was already reduced that it is already identical to water. An arboreal species of frog, Philautus sp. was found living inside the trap of N. saranganiensis (Fig. 5). There were only few insects (mostly ants) were found inside the pitcher plant and almost all of the insects were already digested. However, there was evidence that there were large flying insects around the area due to the presence of a single insect wing attached to the outer surface of the pitcher body.
Table 1. Descriptions of the peristome of N. saranganiensis shape variation in different relative warps.

Table 2. Descriptions of the pitcher body of N. saranganiensis shape variation in different relative warps.

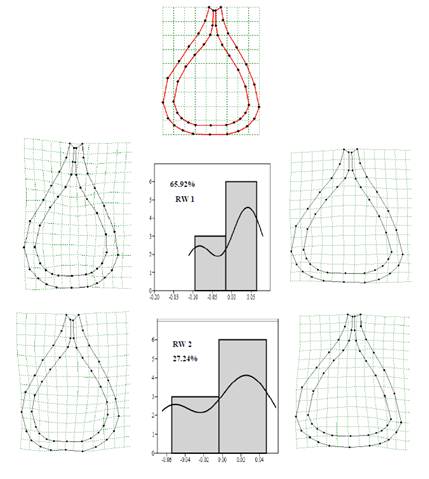
Fig. 6. Graphical illustrations of the relative warps, grid deformations and histograms of the peristome shape of N. saranganiensis.
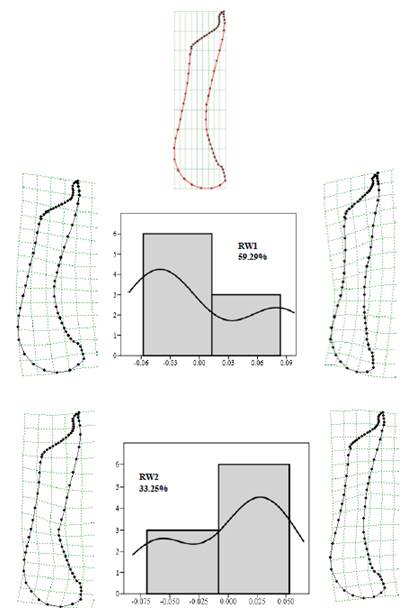
Fig. 7. Graphical illustrations of the relative warps, grid deformations and histograms of the pitcher body shape of N. saranganiensis.
Results on the relative warp analysis had already provided detailed quantitative and qualitative descriptions on the pitcher morphology of N. saranganiensis. Qualitative descriptions based on observations of the external and internal morphology of N. saranganiensis, habitat descriptions, micro-climate measurements and putative macrofaunal associates were determined in this study. With these obtained data, the possible functional morphology of N. saranganiensis pitcher was determined. Findings of the study suggest that the plant relied on its slippery and waxy zone and it specializes on prey retention. Following the criteria created by Moran et al. [35] on grouping species of Nepenthes according to its morphological characteristics and its functional morphology, N. saranganiensis was classified into the "Dry syndrome" group or category. Possible mutualistic activities may have been exhibited by the tree frog Philautus sp. and the pitcher plant where the frog was found inside the pitcher plant, unharmed by the pitcher fluid.
4. Discussion
Based from the Relative Warps Analysis on the shapes of the peristome and pitcher body of N. saranganiensis, the study revealed that the most variable part of the peristome were the lateral and basal regions including the entire structure of the pitcher from the slippery zone, transition zone and digestive zone. Histogram distributions have shown that most of the specimens tend to have wider peristomes and bodies when compared to the shape consensus. However, there were certain populations observed to possess some of the opposite traits wherein they had narrow and upright ad slender pitcher bodies.
These morphological differences in the pitcher may be due to the environmental factors that may have stimulated the phenoplastic characteristics of the plant. Since N. saranganiensis grows epiphytically on the upper and middle canopy of trees on a closed canopy mossy forest, leaves of host trees would often limit light penetration in the forest floor. When parts of the plant are constantly being shaded by the leaves of the trees, it would result to an enlarged surface area especially appendages such as the leaves and pitchers in contrast to those that are exposed to sunlight with normal appendage size. These adaptations maybe due to the plants response to environmental changes which they exhibit dynamic environmental responses and express plasticity for a number of morphological adaptations such as organ size and structure [19]. The best example for this is the size of leaves produced in different light availability conditions in which in this case, the modified leaf or the pitcher of N. saranganiensis. Decreased light availability reduces the total number of leaves produced and the next response is to make these individual leaves as large and wide as possible under growth limits to maximize the light capture area [16, 18, 19, 21]. This physiological response is still applicable in pitcher plants because although they capture and digest insects and other organic material in their traps as source of nitrogen and other nutrients and minerals, they still rely on the sun’s light to process these nutrients during photosynthesis.
For the functional morphology of N. saranganiensis, based on the results of the Relative Warp Analysis and qualitative descriptions of the external morphology of the pitchers, N. saranganiensis had a prominent and developed slippery zone and narrow peristome in relation to its body size. Moran et al. [35] provided the criteria on classifying the trapping strategies of Nepenthes plants. In this case, N. saranganiensis fits to the dry syndrome category based on its morphological characteristics by having a narrow peristome, developed and prominent slippery zone with very visible wax crystals and a less viscoelastic fluid. These dry syndrome Nepenthes are not climatically limited pertaining to their habitat preference wherein most species in this group are predominantly found in dry, warm and non-humid lowlands and some species can be also seen in cold, wet or per humid and highly elevated areas [35]. This also explains the distribution and habitat of N. saranganiensis. In addition, their traps are both effective in these conditions since they specialize both in trapping and retaining their prey where it is very visible on the pitcher body shape observed in the results of Geometric morphometric Analysis and the observed external morphology of N. saranganiensis wherein the globose digestive zone and upright and slender slippery zone lined with slippery wax effectively prevent insects on climbing out of the pitcher. This aspect was investigated by Benz et al. [22] on N. alata where the shape of the digestive zone played an important role on retaining the trapped insects when they tried to climb on the angular surface of the digestive zone due to the contamination of digestive fluids on the insect’s adhesive pads, rendering it useless. They are furthermore hindered by the waxy slippery zone of the pitcher plant even when their adhesive pads have already been dried out by rubbing their feet to their head and antennae, the adhesive pads were still rendered useless due to the waxy lining of the slippery zone of the pitcher where it detaches easily and coating the insect’s foot with wax. Based on the descriptions of McPherson and Amoroso [26] on N. saranganiensis, the plant’s pitchers closely resemble the pitchers of N. alata except to its reduced fringed wings. This morphological resemblance can be furthermore justified by Cheek and Jebb [27] wherein N. saranganiensis was classified under the N. alata species complex. Thus, the observations of Benz et al. [22] on N. alata can be formidably applicable on N. saranganiensis.
The presence of macrofaunal associates were also observed during the field work where insect prey and a possible amphibian symbiont were found inside the pitchers of N. saranganinensis. In montane forests, flying insects are more likely to be observed with some ant species being trapped in the pitchers of highland Nepenthes which were observed on study of Chin et al. [13] Moran et al., [35] and Bauer et al. [3]. As for the Philautus sp. found inside the pitcher plant, it may have used the pitcher plant as its home and hunting or trapping area because insects would be attracted to the plant with its extrafloral nectaries, and it would be much easier for them to catch the insects trapped inside the pitchers or insects hovering or crawling nearby the pitchers. It would only require less energy and effort for the frog to expend on obtaining food. There is a great likelihood that the frog would defecate inside the pitcher after a satisfying and seemingly effortless meal which in return, the pitcher plant may utilize the frog’s feces as a source of nitrogen and other essential nutrients. Hua and Li [36], also observed frogs inside the pitchers of N. mirabilis Druce where they wait inside the pitchers to ambush prey and easily hop in and out from the pitchers. This symbiotic relationship is also true to some species of Nepenthes such as N. rajah where Chin et al., [13] observed the symbiotic relationship of the plant and mountain shrew (Tupaia montana)
5. Conclusion
Land-mark based geometric analysis on the shapes of the peristome and pitcher body of N. saranganiensis revealed that the most variable part of the peristome were the lateral and basal regions including the entire structure of the pitcher from the slippery zone, transition zone and digestive zone. Relative warp analysis likewise provided the quantitative description of peristome shape and body pitcher body of Nepenthes. Based from analysis, results suggest that most of the specimens tend to have wider peristomes and bodies when compared to the shape consensus. Putative functional morphological features of N. saranganiensis exhibit a prominent and developed slippery zone and narrow peristome in relation to its body size. Such adaptation primarily fits within the "Dry Syndrome" trapping strategy as this species mainly specialized in prey retention due to the presence of wax in the slippery zone thereby preventing the trapped insects from escaping the pitcher.
Acknowledgement
The authors are grateful to Dr. Chris Rey Lituañas and Prof. Jennifer Opiso of the Biology Department of Central Mindanao University for reviewing the manuscript, Prof. Muhmin Michael Manting of Mindanao State University- Iligan Institute of Technology for the technical assistance and helpful suggestions and lastly, Mr. Wally Suarez for taxonomic confirmation of the Nepenthes specimen.
References